单分子逻辑开关概述
单分子逻辑开关概述
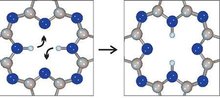
右图所示为单分子逻辑开关整个开关的过程,两个氢原子位于分子中央的一个空洞内。当电压脉冲注入时,两个氢原子变换位置,如左图所示。开关不会改变任何中心空洞外部的分子结构。
通过引导一个电压脉冲通过分子的上下两个尖端,在邻近的分子(分子中心的白色物体)内的两个氢原子改变位置,靠电力控制整个分子的开关。这就构成了一个基础逻辑门,
计算机芯片内的开关与光开关的作用方式一样,用来打开和关闭电子流并将它们放到一起,构成逻辑门,即构成计算机处理器的电路。开关尺寸越小,电路尺寸也就相应的小,从而有可能将更多的电路集成到一个处理器上,同时还可以提高速度和性能。
这种分子开关的出现使得制造尺寸超小、但是速度堪比超级计算机的芯片成为可能;甚至还有可能产生只有一丁点灰尘那么大或可以放到针尖上的计算机芯片。
单分子逻辑开关造价信息
单分子逻辑开关概述常见问题
-
简单理解就是控制电流断开或者闭合的开关,主要的参考可为温度、压力、流速、液位。SOR 是品牌,生产商是美国的。主要是膜片活塞机械式的。产品质量不错。
-
主要有这几种: 1、户内空气绝缘开关柜(UniGearZS1 ,Ungear ZVC, Unigear Z3.2 ,Unigear 550) 2﹑户内气体绝缘开关柜(ZX2,ZX0,ZX1.2,ZX1...
-
1.列清单的时候有逻辑顺序吗,怎样才能做到不漏项呢? 没有严格说明,一般编号应该从小到大,倒过来说明编制人在编制清单中有漏项,后补的项目。 2.精装修东西那么多,感觉很杂乱,前辈可有心得? 我做了20...
单分子逻辑开关概述文献

 基于双控荧光开关的NOR和OR分子逻辑门
基于双控荧光开关的NOR和OR分子逻辑门
合成了L-精氨酸蒽衍生物1,考察了碱土及过渡金属离子对主体分子1荧光光谱的影响,结果发现,在中性水溶液条件下只有Cu2+能有效地猝灭其荧光.另外,通过OH-/H+和Cu2+/乙二胺四乙酸(EDTA)均能对化合物1的荧光强度进行可逆性调控.在此基础上,我们以化合物1水溶液作为起始状态,以OH-和Cu2+为两化学输入,构建了一个"或非"(NOR)分子逻辑门;以1-Cu2+水溶液体系作为起始状态,以H+和EDTA为两化学输入,构建了一个"或"(OR)分子逻辑门.
 数字逻辑第1章开关理论基础
数字逻辑第1章开关理论基础
数字逻辑第1章开关理论基础
分子开关的定义
molecular switches,通过激活机制或失活机制精确控制细胞内一系列信号传递的级联反应的蛋白质。
分子开关的分类
细胞内信号传递作为分子开关的蛋白质可分两类:一类开关蛋白(switch protein)的活性由蛋白激酶使之磷酸化而开启,由蛋白磷酸酯酶使之去磷酸化而关闭,许多由可逆磷酸化控制的开关蛋白是蛋白激酶本身,在细胞内构成信号传递的磷酸化级联反应;另一类主要开关蛋白由GTP结合蛋白组成,结合GTP而活化,结合GDP而失活。
英国朴次茅斯大学的科学家指出:有一天,这些微小的开关很可能形成微电子回路的基础,帮助人们更加快速和准确地解决DNA的排序问题。
这种新型的分子开关是通过固定在DNA上的微小金属珠的摆动来拉动一根DNA链的(如图)。双螺旋链的一端被附着在一个微芯片的微小通道上,DNA的另一端安放金属珠。这些金属珠只有1微米宽,也就是一根人头发丝直径的1/50。
珠子是顺磁性的,即在磁场中其行为就像其本身是一块磁铁。其结果,小珠子可以被拽向磁场,使DNA链立起。
接下去科学家将把可在 DNA上转动的发动机安装到DNA链上。这个发动机是一种自然产生的蛋白质,叫做限制修饰酶(restriction-modification enzyme),燃料是三磷酸腺苷(ATP)。由糖、磷酸盐和核苷碱基腺嘌呤组成的分子能够为肌肉和其他生物组分供能,蛋白质只是结合在DNA链的特定位点上。
所有的DNA都由四种类型的核苷碱基组成,即腺嘌呤(A),鸟嘌呤 (G),胞核嘧啶(C)和胸腺嘧啶(T)。朴次茅斯大学的分子生物技术专家弗门(Keith Firman)说:"因为蛋白质发动机只能允许自己附着在DNA碱基的某些特定顺序上。这就使得科学家得以准确地控制它在DNA上的位置。"
当ATP燃料被添加在开关周围时,发动机会拉动DNA,在它下面绕圈,一直到它抵达金属珠。然后金属珠会撞上发动机,就像一根多节的绳子卡到了一个滑轮里一般。
当发动机耗尽了燃料的时候,它就脱离开DNA链,磁场于是重新将DNA拉紧。
正像任何磁性材料,顺磁的珠子在通过磁场的时候也将产生电信号。为了侦察到这种微弱的信号,科学家在DNA的通道基部放置了一些叫做"霍尔效应传感器"(Hall Effect sensor)的灵敏传感器。这些传感器可以感受到磁体的移动。
所以,在添加了燃料的时候,这个微装置就能发出开关的控制信号。弗门说:"叫它开关,因为它是被ATP燃料激活的,是用在发动机上的。当被激活时,它就会打开电子装置,而当它没有被激活时,就关闭了。
弗门还指出:未来,人们有可能制造出可以控制纳米管中流动材料的珠子。直接拉动DNA 链,让珠子移动进入纳米管,可以封锁住液体的流动;直接用开关释放DNA ,让金属珠移向磁场,就可以让液体再次通过。
对于通过细胞表面受体所介导的信号通路而言,除受体本身作为离子通道而起效应器作用的情况之外,其他的信号通路首先要完成配体结合所诱发的信号跨膜转导,随之要通过细胞内信号分子(包括第二信使)完成信号的逐级放大和终止。在细胞内一系列信号传递的级联反应中,必须有正、负两种相辅相成的反馈机制进行精确控制, 因此分子开关(molecular switches)的作用举足轻重,即对每一步反应既要求有激活机制又必然要求有相应的失活机制,而且二者对系统的功能同等重要。细胞内信号传递作为分子开关的蛋白质可分两类:一类开关蛋白(switch protein)的活性由蛋白激酶使之磷酸化而关闭,由蛋白磷酸酯酶使之去磷酸化而开启,许多由可逆磷酸化控制的开关蛋白是蛋白激酶本身,在细胞内构成信号传递的磷酸化级联反应;另一类主要开关蛋白由GTP结合蛋白组成,结合GTP而活化,结合GDP而失活。
任何机器都需要开关控制启动或关闭,麻雀虽小、五脏俱全的纳米机器也一样有和它们匹配的微小开关。分子开关(molecular switches)或者叫摩尔开关(mol.switches)就是这样一种能够控制比它们本身稍大的纳米装置的精巧结构。它们可以在纳米世界中发送信息遥控正常大小的传感器。
单分子逻辑开关相关推荐
- 相关百科
- 相关知识
- 相关专栏
- 感应电机智能调速
- 磁力夹具
- 有机硅扩散泵油
- 多功能真空实验仪
- 电工学实验与测量
- 电动机的起动控制与变频调速
- 朗能NB83Q/1
- 西蒙S51011BY
- 真空冷却
- 真空净油机
- 优腾电工
- 润枫嘉尚
- 声光双控延时照明灯
- 施耐德E8433/2
- 施耐德EV54
- 华燕118D-32
- 分形DGS单元结构设计及其在微带电路中的应用
- 中国机械工程学会授予材料热处理工程师资格人员名单
- 基于超磁致伸缩效应的新型液压高速开关阀
- 水利管理单位信访工作
- 中华人民共和国国家标准建设工程工程量清单计价规范
- 新型的低压成套开关设备GGD型低压配电柜
- 应用清单招标与经评审的最低投标价法评标的几点思考
- 招投标书—转轮除湿机方案概述技术标商务标标书模板
- 招标师继续教育-WTO政府采购协议(GPA)概述
- 有关建设工程施工安全涉及常用法律法规标准规范清单
- 保护装置BUCK开关电源的PCB优化设计方法
- 云南省氯碱PVC行业清洁生产合格单位评价指标体系
- 新型空间梁单元及其在梁杆结构稳定分析中的应用
- 中国会计学会优秀会计学术论文获奖名单
- 云南省机关事业单位编制外人员劳动合同书(修改后)
- 新型陶瓷化高分子复合耐火硅橡胶耐火电缆的研制
最新词条
安徽省政采项目管理咨询有限公司
数字景枫科技发展(南京)有限公司
怀化市人民政府电子政务管理办公室
河北省高速公路京德临时筹建处
中石化华东石油工程有限公司工程技术分公司
手持无线POS机
广东合正采购招标有限公司
上海城建信息科技有限公司
甘肃鑫禾国际招标有限公司
烧结金属材料
齿轮计量泵
广州采阳招标代理有限公司河源分公司
高铝碳化硅砖
博洛尼智能科技(青岛)有限公司
烧结刚玉砖
深圳市东海国际招标有限公司
搭建香蕉育苗大棚
SF计量单位
福建省中亿通招标咨询有限公司
泛海三江
威海鼠尾草
Excel 数据处理与分析应用大全
广东国咨招标有限公司
甘肃中泰博瑞工程项目管理咨询有限公司
山东创盈项目管理有限公司
当代建筑大师
广西北缆电缆有限公司
拆边机
大山槟榔
上海地铁维护保障有限公司通号分公司
甘肃中维国际招标有限公司
舌花雏菊
华润燃气(上海)有限公司
湖北鑫宇阳光工程咨询有限公司
GB8163标准无缝钢管
中国石油炼化工程建设项目部
韶关市优采招标代理有限公司
莎草目
建设部关于开展城市规划动态监测工作的通知
电梯平层准确度
广州利好来电气有限公司
四川中泽盛世招标代理有限公司